补硒能提高嘌呤代谢吗(代谢能补嘌呤提高硒吗)
硒通过硒蛋白发挥生物学作用
硒进入机体后,与体内氨基酸共价键结合形成硒蛋白来表现其生物学作用,调控机体 的自由基代谢、抗氧化功能、免疫功能、生殖功能、细胞凋亡和激素分泌等。所以说,硒 的功能是通过硒蛋白来表现的(张在香,等,2000)。
到目前为止,在硒蛋白中只发现两种含硒的氨基酸(图 7-1):一种由硒原子取代了 半胱氨酸中的硫原子形成的硒代半胱氨酸(Sec)。构成人体蛋白质的基本氨基酸有 20 种, 因此人们又将硒代半胱氨酸称为生物体内的第二十一种氨基酸。另一种是由硒原子取代了 甲硫氨酸中的硫原子形成的硒代甲硫氨酸(SeMet)。有的文献称“硒代甲硫氨酸”为“硒 甲硫氨酸”。
这两种含硒氨基酸统称为“硒代氨基酸”,它们都参与多种硒蛋白的合成。
硒蛋白合成是一个非常复杂的生物学过程。原核生物中硒蛋白的合成较简单,而真核 生物中硒蛋白合成过程十分复杂。在这个过程中,硒代甲硫氨酸可以替代甲硫氨酸掺入到蛋白质分子中,而且这种替代是非特异性的,当生物体内甲硫氨酸不足时,硒代甲硫氨酸 可以被诱导而替代甲硫氨酸参与硒蛋白的合成 ,取代后的硒蛋白活性不会受到影响(程 天德,吴永尧,2004)。而硒代半胱氨酸掺入蛋白质是由密码子 UGA 介导的翻译过程(徐 辉碧,等,2002)。即在含有硒代半胱氨酸的蛋白质合成过程中,先由信使核糖核酸(mRNA) 上的 UGA 识别与硒代半胱氨酸结合的转运核糖核酸(tRNA),再经过复杂的翻译过程, 将它掺入到蛋白质中,形成含有硒代半胱氨酸的蛋白质而发挥其生物学作用。UGA 是硒 代半胱氨酸的密码子,它的发现,为硒的生物化学研究开辟了新的天地(王海宏,谢忠忱, 2003)。
① tRNA[Ser]Sec:在原核生物合成硒蛋白过程中,有一种 tRNA 既可与丝氨酸(Ser)结合,也可与硒代半胱氨酸(Sec) 连接,这种特殊的 tRNA 是 SelC 基因的产物。为了区别于其他 tRNA,有学者用“tRNA[Ser]Sec”表示,现在也有用“tRNASec” 表示的。
目前发现人硒蛋白有 25 种:5 种 GPx(GPx1、GPx2、GPx3、GPx4 和 GPx6), GPx1 ~ GPx4 存在于所有哺乳动物中,GPx6 在多个物种中是硒蛋白,但在少数物种中是含 Cys的蛋白质;3种甲状腺激素脱碘酶(ID1、ID2和ID3);3种硫氧还蛋白还原酶(TXNRD); 硒磷酸合成酶(或硒代磷酸酶 SPS2);以及硒蛋白 F(原名为 15kDa 硒蛋白)、硒蛋白 H、 硒蛋白 I、硒蛋白 K、硒蛋白 M、硒蛋白 N、硒蛋白 S、硒蛋白 O、硒蛋白 P、甲硫氨酸 亚砜还原酶 B(原名为硒蛋白 R)、硒蛋白 T、硒蛋白 V 和硒蛋白 W(哈特菲尔德,等, 2018)。
研究发现内质网中存在 7 种硒蛋白,分别是 2 型脱碘酶(ID2)、15kDa 硒蛋白、硒 蛋白 M 、硒蛋白 T 、硒蛋白 K 、硒蛋白 S 和硒蛋白 N 。内质网是哺乳动物细胞内重要的 细胞器和最大的膜网络结构,位于细胞核附近。多种生理或病理情况会引起内质网应激, 现发现内质网应激与多种疾病有关。刘红梅等(2014)、毋瑞朋等(2014)将这 7 种硒蛋 白在内质网中的功能、在内质网应激中的作用也做了介绍,并指出,目前对这 7 种内质网 硒蛋白的结构与功能研究很有限。加强对它们的结构、功能及在内质网应激应答中的作用 和机理研究,对于全面了解硒的生物功能、阐明内质网应激相关疾病的病理具有重要的科 学意义。
下面对这些人硒蛋白做一些简要介绍:

(1)GPx:该酶于 1957 年由 Mills 和 Randall 首先发现,直到 1973 年才由 Rotruck 证实它是一类含硒酶。这是人类最早发现的硒蛋白。
低硒为什么对机体健康有如此广泛的影响? GPx 的发现,初步揭开了这个生命现象 的疑团。
研究发现,GPx 是哺乳动物体内最丰富的硒蛋白。每一 GPx 分子有 4 个硒原子与之 结合,形成酶分子的活性中心①发挥作用。硒的含量变化将直接影响到该酶的活性,所以, GPx 也叫硒依赖酶。在代谢中,硒参与催化谷胱甘肽(GSH)还原体内的过氧化物(如有 机氢过氧化物 ROOH、过氧化氢 H2O2 等)的反应:

这个反应将代谢过程中生成的有毒性的过氧化物还原成无毒的物质,从而消除过氧化物对正常组织的破坏作用(黄开勋,等,2009)。故 GPx 是机体非常重要的过氧化物分解酶, 能有效保护细胞及其他敏感生物分子(如DNA、蛋白质、脂质体等)免受氧自由基的损伤(程 彩虹,等,2013)。
解释:① 酶分子活性中心,是指酶分子结构中能发生空间结构改变的那部分结构。它能和底物特异性结合并将底物转化 为产物,生物化学将这一区域称为酶的活性中心。
目前,国内外文献对 GPx 报道比较零散。华中科技大学甘璐教授比较全面和系 统地概括了该酶的类型与功能(黄开勋,等,2009)(参见表 7-1)。
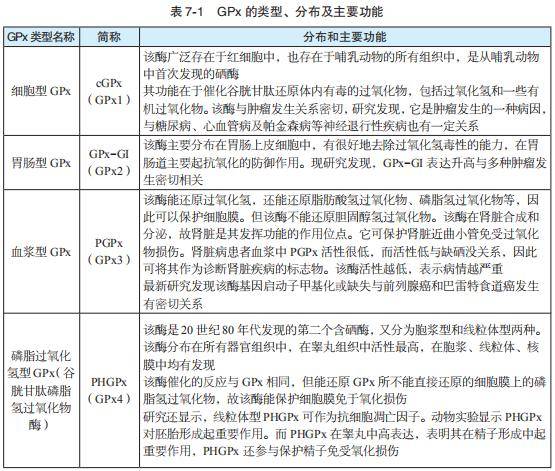
但Brigelius-Flohé和Kipp两位学者(2009)认为,并不是所有GPx都有防癌、抗癌作用, GPx2 在肿瘤的起始阶段具有抗炎抗肿瘤作用,在肿瘤形成后,又支持肿瘤生长,所以, GPx2 可能具有抗癌、致癌的双重性。
(2) 甲状腺激素脱碘酶:刘红梅教授在《硒的化学、生物化学及其在生命科学中的 应用》中对该酶的合成、功能等有详细描述。这里只做一简要介绍:
含硒的脱碘酶又称碘甲腺原氨酸脱碘酶(ID),早已被科学家所知。该酶分为三种类 型:Ⅰ型(ID1)、Ⅱ型(ID2)、Ⅲ型(ID3)。
脱碘酶的功能是调节甲状腺素的代谢:将活性低的 T4 转化为活性高的 3,5,3′- 三碘甲状腺氨酸(T3),再将活性高的 T3 转化为无活性的 rT3、3,3′- T2。硒是脱碘酶的必需 组成成分(夏弈明,1993)(参见“12.3现代科学证明甲状腺病还与缺硒密切相关”)。
同时,该酶对胚胎发育也至关重要,在胚胎、新生儿和儿童期介导细胞增殖和分化。先天 性甲状腺功能减退症主要是甲状腺激素合成和分泌的缺陷导致甲状腺激素缺乏,进而引起 呆小症,表现为身材矮小,精神发育迟缓(郭翠翠,2014)。
ID2是目前研究得最清楚的内质网硒蛋白。它能够催化3,5,3’,5′-四碘甲腺原氨酸(T4) 转化为有生物活性的 T3。在人体内,ID2 是垂体前叶、甲状腺、骨骼、心肌、胎盘和棕色 脂肪组织中 T3 的重要来源,是成人中枢神经系统中唯一的 5′- 脱碘酶。
(3)硫氧还蛋白还原酶(TrxR,TXNRD):TrxR 主要存在于胞浆、线粒体、肝脏 等中,其中 TrxR3 在睾丸呈高表达。该酶具有多种生物活性,能调节机体的氧化还原、 防御氧化损伤、调节细胞生长与增殖、参与信号转导和调节免疫应答等(黄开勋,等, 2009)。
据刘琼等(2009)的综述,TrxR 对肿瘤的发生具有双重效应。在肿瘤发生之前,TrxR 能抗氧化损伤、保护细胞、防止癌变。然而,一旦肿瘤形成,肿瘤的发生依赖于 Trx/TrxR 系统脱氧核糖核酸的补给,TrxR 就具有促肿瘤形成活性,肿瘤细胞中 TrxR 成倍增长。因此, TrxR 在与癌症的关系上具有双重性,既能提高 p53 蛋白①的稳定性,抑制肿瘤增殖,也能通 过硫氧还蛋白与凋亡信号调节激酶 1(ASK1)结合,抑制肿瘤细胞凋亡。TrxR 成了肿瘤的 特异性标志物之一,是癌症治疗的一个新靶点。北京大学曾慧慧教授研究团队以该酶为靶标, 设计和合成了新型含硒抗癌药物乙烷硒啉(BBSKE)。该药物对多种肿瘤细胞中 TrxR 起到 有效的抑制作用,目前正在进行二期临床试验(陈宝泉,等,2011)。
TrxR 还与艾滋病、自身免疫性疾病、感染性疾病和遗传性疾病的发生和发展密切相关, 是潜在的干预治疗靶点(黄开勋,等,2009)。
(4)硒蛋白 P(SelP,SELENOP):SelP 是 Burk 等发现、Tappel 命名的。1987 年 Burk 首次用单克隆抗体分离纯化了大鼠血浆的 SelP。1993 年又从人的血浆中分离纯化了 人的 SelP,故 SelP 亦叫血浆蛋白,是血浆中最丰富的硒蛋白,在脑、肝和睾丸中高表达, 在调节体内硒平衡和运输中起关键作用。动物实验显示,SelP 为一种硒转运蛋白,为大脑 和睾丸组织转运硒(张在香,等,1998;夏弈明,等,2011;潘利斌,等,2017)。它还 是一种氧化还原酶,起着抗氧化作用;对重金属毒性有重要拮抗作用;对精子产生和雄性 生殖具有重要作用等等等。
目前,除少数硒蛋白外,人们对大多数硒蛋白的生物学功能以及表达和调控细节还不 完全了解,甚至很不了解,许多还有待今后去探索。所以,科学工作者对硒蛋白的研究有 着极其广阔的空间(徐辉碧,等,2002;夏弈明,2011)。
继续阅读
- 暂无推荐
